Witaj w sklepie internetowym z suplementami diety premium!
Nasza oferta jest uważnie selekcjonowana pod kątem jakości i skuteczności, dzięki czemu uzupełnianie nawet poważnych niedoborów składników odżywczych staje się całkowicie skuteczne i bezpieczne.
Znajdziesz u nas m.in. najlepszą na świecie liposomalną witaminę C i liposomalny glutation jak inne innowacyjne i unikalne na rynku suplementy diety idealne dla ŚWIADOMYCH JAKOŚCI ODBIORCÓW

>>NEUTRIENT™
nowa europejska marka suplementów diety tworzona przez praktyków. Unikalne połączenia składników odżywczych

>>ALTRIENT LIVON LABS™
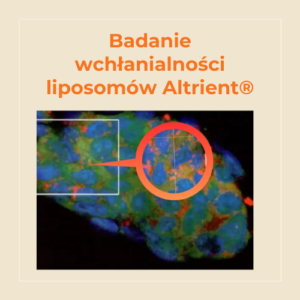
Pierwsza na świecie linia liposomalnych suplementów diety. Od 2004 roku niezmiennie wiodąca pod względem jakości i efektywności.

>>ResultsRNA™
unikalne na rynku amerykańskie suplementy i kosmetyki o wyjątkowych właściwościach absorpcyjnych
Bestsellery


Propozycje Według Twoich Potrzeb













OUTLET
Krótkie daty – super rabaty! Ale nie tylko. Czasem otrze się lub zgniecie opakowanie a my je przecenimy. Zajrzyj!

WIĘCEJ PROPOZYCJI
dla uzupełnienia planu suplementacyjnego wyszukujemy na rynku preparaty, które spełniają wysokie standardy jakościowe i absorpcyjne, oraz kosmetyki bezpieczne dla skóry i środowiska

ZESTAWY -10%
w oparciu o naszą wiedzę i doświadczenia stworzyliśmy Zestawy tak, aby jak najlepiej służyły zdrowiu naszych Klientów.